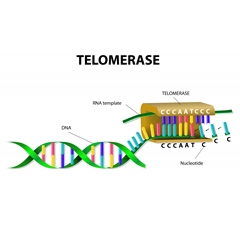
寡核苷酸类端粒酶抑制剂通过与端粒酶功能性 RNA互补序列功能性 RNA结合,抑制端粒酶的功能,实现抑制肿瘤生长。
1938年,Hermann Muller注意到染色体的末端对X射诱导突变,具有抵抗能力,并把染色体的末端命名为端粒。
1972年,James Watson提出DNA末端复制的问题,由于DNA亲代的3ʹ端需要几个自由碱基与RNA引物结合来引导DNA复制,结果造成亲代模板3ʹ端不能实现完全复制,DNA每一次复制,DNA链都会缩短,导致末端遗传信息的丢失(J D Watson., 1972)。
1978年,Blackburn和Gall发现嗜热四膜虫(Tetrahymena Thermophila)端粒中存在20至70次TTGGGG重复序列(BE H Blackburn., 1978)。
1988年,R K Moyzis等人确定人端粒中存在类似的重复序列TTAGGG(R K Moyzis., 1988)
1985年,Greider和Blackburn提出存在一种机制能够将重复序列添加到DNA的末端(C W Greider., 1985)。
1987年,Greider和Blackbur鉴定出能够将重复序列添加到DNA3ʹ端,延长端粒的端粒酶(C W Greider., 1987)端粒酶属于RNA依赖性DNA聚合酶,主要由功能性RNA和逆转录酶两部分构成。功能性RNA 由TERC基因编码被称为TERC或TR,逆转录酶由TERT基因编码,被称为TERT。此外还包括TCAB1和H/ACA复合体来协助组装TERC(George E Ghanim., 2024)。
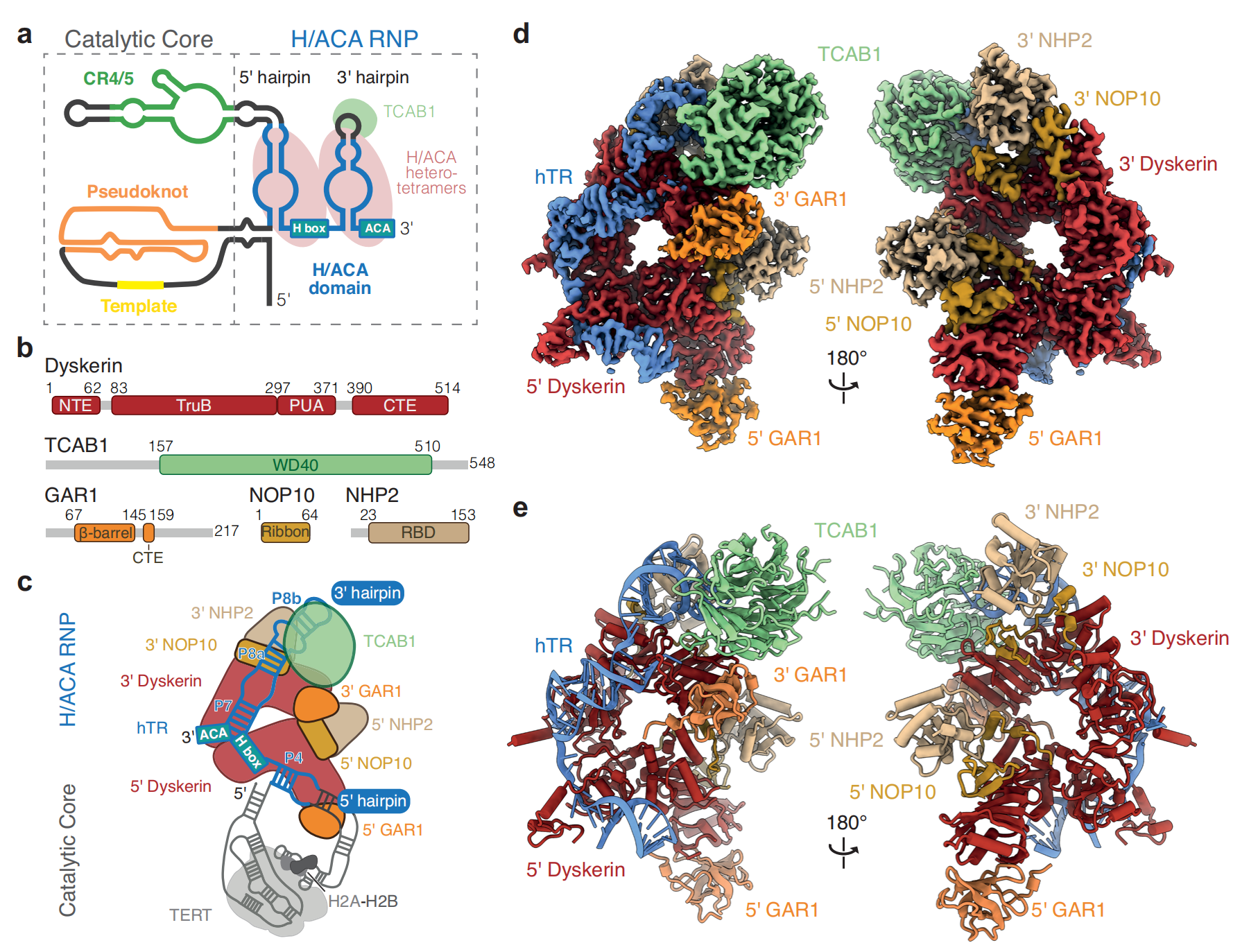
1994年,C M Counter等人的研究显示,卵巢上皮癌细胞能够表达端粒酶,而非癌卵巢上皮细胞不能表达端粒酶。提示端粒酶在癌细胞永生化中发生重要作用(C M Counter., 1994)。
抑制端粒酶,可供考虑策略是是开发能够与TERC互补的寡核苷酸。由于非修饰寡核苷酸在体内代谢迅速,并且难以透过细胞膜,因此需要进行修饰,改善在体内的分部和延长作用时间。对于寡核苷酸修饰,主要包括核糖部分和磷酸酯骨架部分。核糖修饰除了能够增加寡核苷酸的亲脂性,还会使寡核苷酸能够耐受核酸酶的降解。对磷酸酯骨架部分进行修饰,可以增加寡核苷酸的脂溶性,延长作用时间,并使用寡核苷酸更容易通过细胞膜(Brittney-Shea Herbert., 2005)。
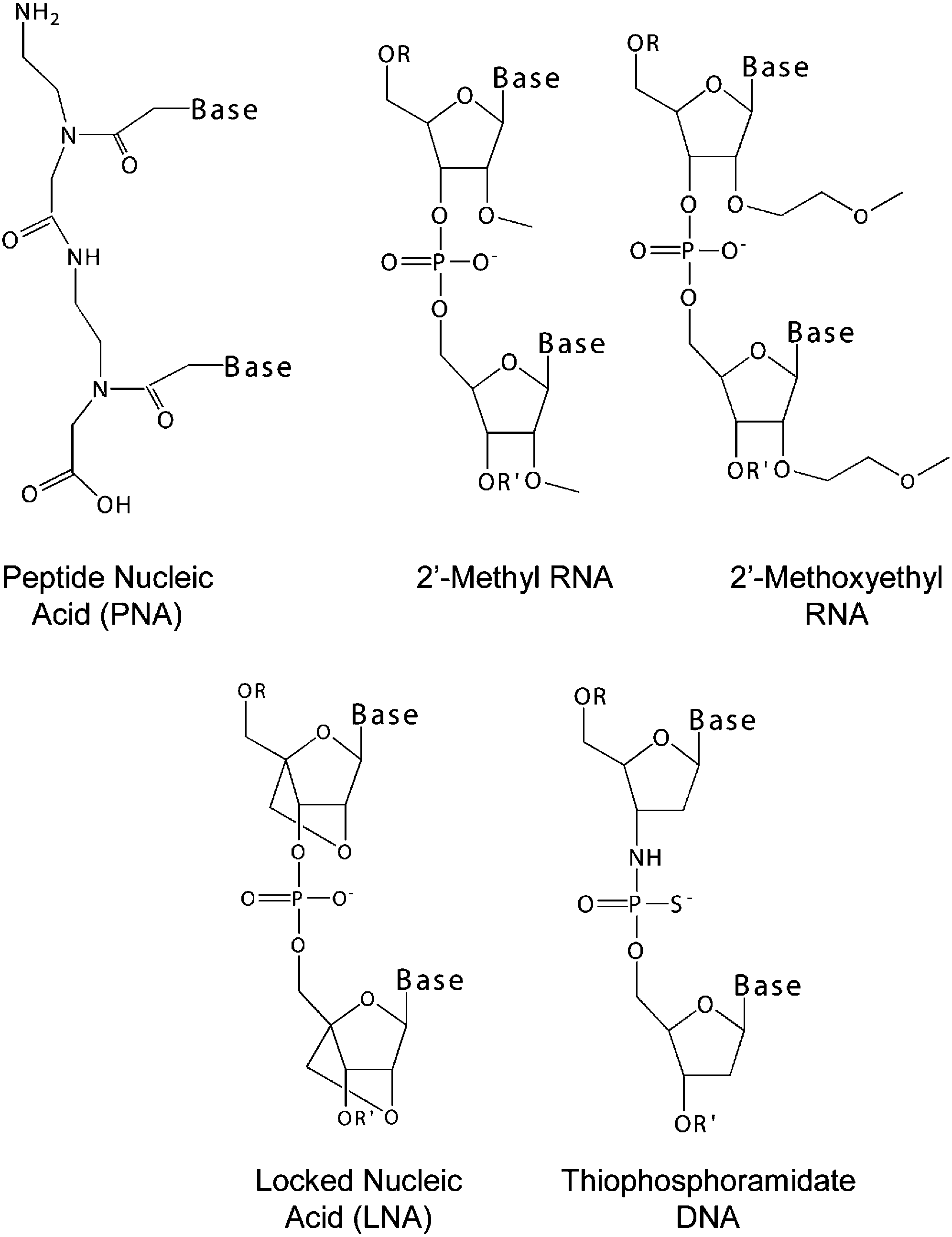
1996年,J C Norton使用肽核苷酸片断,实现抑制端粒酶的活性(J C Norton., 1996)。
2001年,S Gryaznov引入硫代磷酰胺替代寡核苷酸的磷酸酯骨架,实现抑制端粒酶的活性(S Gryaznov., 2001)。
2003年,Zhi Chen等人使用核糖2ʹ位由2ʹ-O-甲氧基乙基进行修饰的寡核苷酸,来抑制端粒酶,实现阻止肿瘤生长(Zhi Chen., 2003)。
免责声明:相关信息仅限药物研发参考使用,本网站不保证信息真实和准确!
关注“药研苑”公众号,查看前景解析。

|
推荐阅读: ・2025年1-3季度口崩片市场哪个强? ・2025年1-3季度口溶膜制剂哪家强? ・免疫检查点抑制剂,谁将成为下一个王者? ・质子泵相关抑酸类药物市场即将回暖 ・中药1类新药距离封神还有多远 |
|
“药品营销避坑”试读: ・药品立项需要注意什么(一) ・药品生命周期管理(一) ・改剂型,口服液体制剂“金矿”还是“陷阱 |
我们提供如下咨询服务:药品信息发布、药品立项、市场前景分析、医院及药品零售市场分析、药品市场调研及制定推广策略、国外药品引进、国内批文转让、上市前后临床试验设计、药品彩页及主图设计、药品推广PPT制作。您可以关注公众号“药研苑”后,在主页面发送消息,咨询相关服务。


横切线®为注册商标
Copyright 2020 横切线®药研苑 备案号:粤ICP备18041379号-3