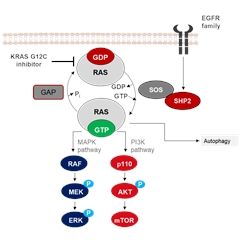
KRASC抑制剂能够抑制KRAS的过度激活。
1964年Jennifer Harvey发现与既往使用的莫罗尼白血病病毒(Moloney's Leukaemia Virus,MLV)不同,其从一只白血病大鼠中提取制成的莫罗尼白血病病毒制剂,诱导新生BALB / c小鼠发生肉瘤(J J HARVEY., 1964)。
1967年Kirsten等人使用小鼠骨髓成红血细胞增多症病毒诱导大鼠出现淋巴瘤,并进一步诱导出肉瘤(W H Kirsten., 1967)
1973年Scolnick等人发现Kirsten的鼠肉瘤病毒(MSV)是Kirsten的白血病病毒杂合大鼠基因,转变而成(Scolnick., 1973)。
1977年Parks从Scolnick Harvey鼠肉瘤病毒(MSV)表达产物中分离出分子量为21000的特殊蛋白质(W P Parks., 1977)。将这种蛋白质被称为P21蛋白,也被称为Harvey RAS。
1979年Shih等人从Kirsten鼠肉瘤病毒表达产物中也鉴定出相类似的P21蛋白(T Y Shih., 1979),也被称为Kirste RAS。
1982年,Parada等人从人类膀胱癌细胞系中分出Harvey RAS基因的同源基团,命名为 HRAS(L F Parada., 1982)。同年Der等人从人类肺癌细胞系系中分出Kirsten RAS基因的同源基团,命名为 KRAS(C J Der., 1982)。
1983年,K Shimizu等人在SK-N-SH人神经母细胞瘤基因中发现第三种RAS,命名为NRAS(1983., K Shimizu)。
1979年,Scolnick等人发现RAS能够与GTP(三磷酸鸟苷)或GDP(二磷酸鸟苷)发生结合(E M Scolnick., 1979)。1984年,Gibbs等人证实RAS是一种三磷酸鸟苷酶(GTPase)(J B Gibbs., 1984)。由于仅含有一条多肽链,RAS属于小GTPase。
小GTPase存在两种状态,“开”和“关”,当RAS与GTP结合时,RAS处于活性状态,激活RAS-RAF-MEK-ERK信号通路,调节细胞周期和增殖。RAS处于活性状态还能够激活PI3K-AKT-mTOR信号通路,调控细胞存活、肿瘤侵袭和转移。
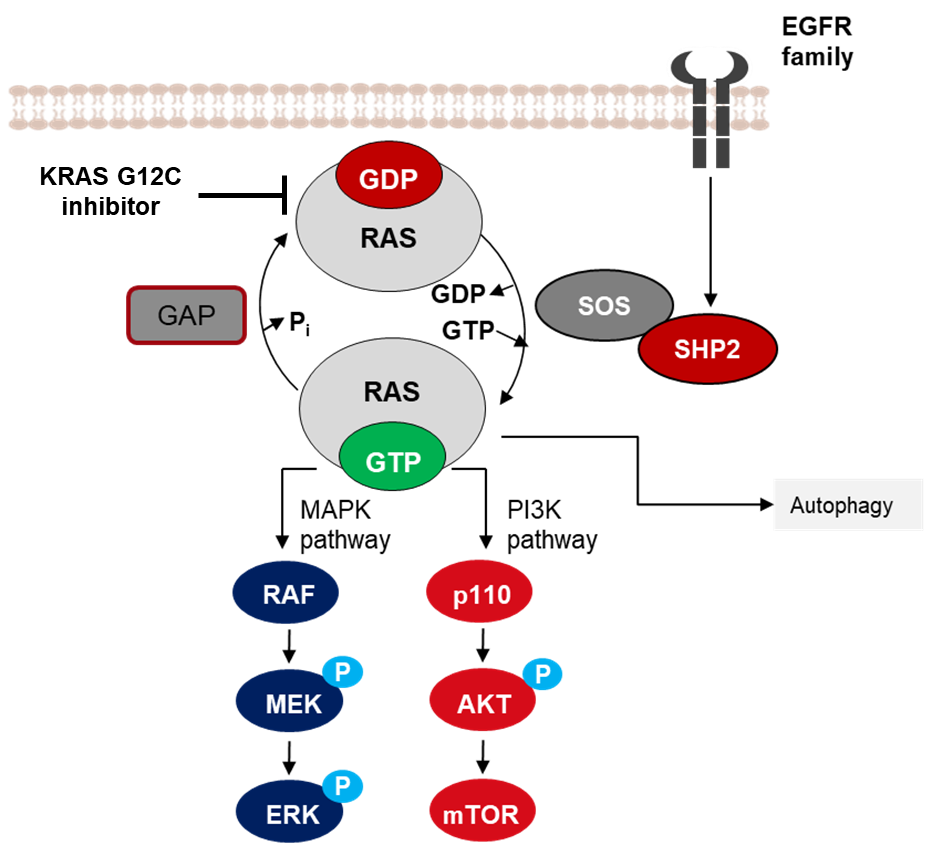
当小GTPase与GDP结合时,GTPase处于失活状态。GTP酶激活蛋白(GAP)能够将GTP水解成GDP,从而使GTPase失活。鸟苷酸核苷酸交换因子(GEF)能够介导GDP与GTP的交换,从而激活小GTPase。与GEF相反,鸟苷酸解离抑制因子(GDI)能够抑制GDP的释放。在GAP、GEF和GDI共同作用下,实现GTPase的精准调控。
1983年Shimizu等人发现人类肺癌细胞系KRAS与Kirsten鼠肉瘤病毒相似,都存在KRAS蛋白12位点突变(K Shimizu., 1983)。甘氨酸突变为半胱氨酸(G12C)。
G12C突变,会导致GDP/GTP交换加速,RAS与GAP结合减少,使KRAS处于过度激活状态。
KRAS的结构包括两个部分,G结构域和高变区(HVR)。

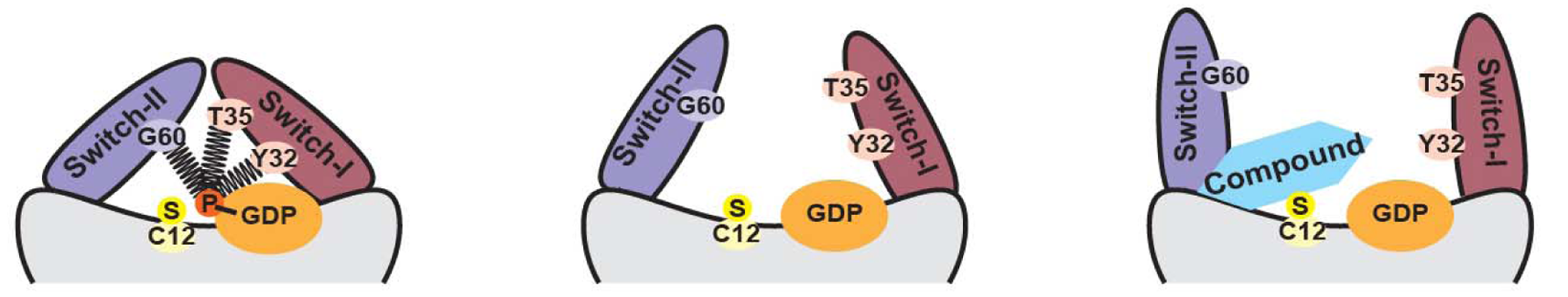
G结构域包括六条β链,组成KRAS结构的核心,在β周围环绕着五个α-螺旋。KRAS的重要功能结构是开关(Switch)区域,即Switch Ⅰ(开关Ⅰ)和Switch Ⅱ(开关Ⅱ)。此区域负责与KRAS的效应蛋白,及KRAS的调节蛋白(如GAP、GEF)结合。
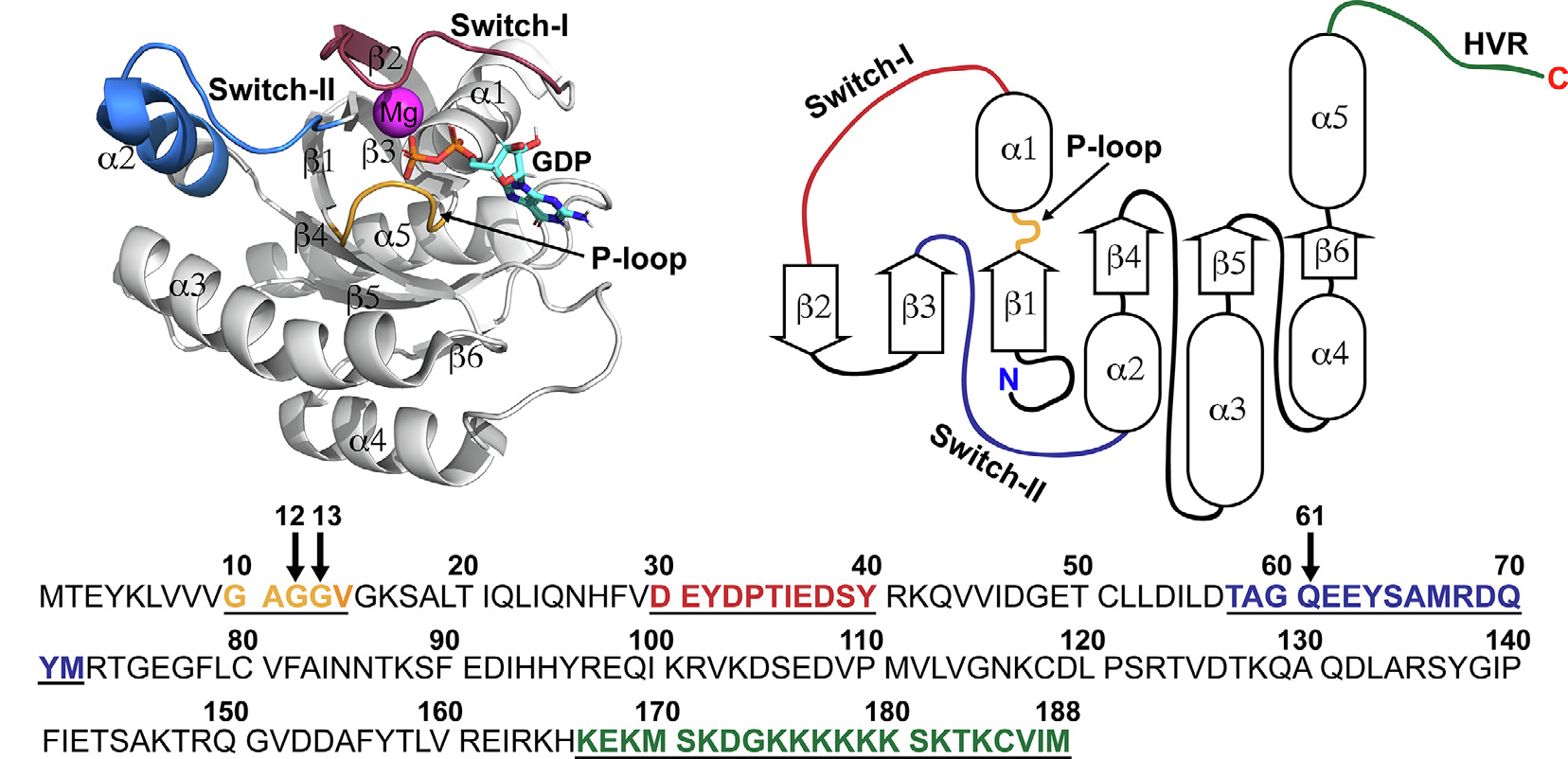
2013年Ostrem等人发现在Switch Ⅱ存在一个用KRAS G12C抑制剂的结合口袋,使KRAS更倾向于与GDP结合(Jonathan M Ostrem., 2013)。
2021年6月20日FDA批准第一个KRAS G12C抑制剂,Amgen安进的Sotorasib(商品名Lumakras)用于治疗KRAS G12C突变的局部晚期或转移性非小细胞肺癌(NSCLC)。
免责声明:相关信息仅限药物研发参考使用,本网站不保证信息真实和准确!
关注“药研苑”公众号,查看前景解析。

|
推荐阅读: ・2025年1-3季度口崩片市场哪个强? ・2025年1-3季度口溶膜制剂哪家强? ・免疫检查点抑制剂,谁将成为下一个王者? ・质子泵相关抑酸类药物市场即将回暖 ・中药1类新药距离封神还有多远 |
|
“药品营销避坑”试读: ・药品立项需要注意什么(一) ・药品生命周期管理(一) ・改剂型,口服液体制剂“金矿”还是“陷阱 |
我们提供如下咨询服务:药品信息发布、药品立项、市场前景分析、医院及药品零售市场分析、药品市场调研及制定推广策略、国外药品引进、国内批文转让、上市前后临床试验设计、药品彩页及主图设计、药品推广PPT制作。您可以关注公众号“药研苑”后,在主页面发送消息,咨询相关服务。


横切线®为注册商标
Copyright 2020 横切线®药研苑 备案号:粤ICP备18041379号-3


